содержание .. 9 10 11 12 ..
Исследование по феногенетике признаков поведения у собак (В.П. Крушинский) - часть 11
ГЕНЕТИКА И ФЕНОГЕНЕТИКА
ПОВЕДЕНИЯ ЖИВОТНЫХ*
Генетические исследования поведения имеют существенное значение
для ряда областей биологии и медицины. Во-первых, они должны быть
той базой, на которой только и могут развиваться некоторые области
физиологии высшей нервной деятельности. Учение об индивидуальных
различиях высшей нервной деятельности (в том числе учение о ее типах)
и выяснение относительной роли врожденных и индивидуально приобре-
тенных компонентов поведения невозможны без генетического анализа.
Вполне понимая это, И.П. Павлов создал в Колтушах лабораторию генети-
ки высшей нервной деятельности.
Во-вторых, генетика дает возможность при помощи скрещиваний
разъединять и соединять в гибридном потомстве те и другие особеннос-
ти поведения с различными морфофизиологическими свойствами орга-
низма и выяснять корреляционные зависимости между теми и другими.
Это открывает новый тонкий метод изучения зависимости формирования
поведения от морфофизиологических свойств организма, который
невозможен при помощи современного хирургического или физиологи-
ческого метода.
В-третьих, изучение генетики поведения имеет большое значение
для ряда проблем эволюционного учения. Генетически обусловленные
особенности поведения животных играют роль в структуре популяций.
Наследственные различия в поведении обусловливают образование
изолированных популяций различной численности, что имеет большое
значение для темпа эволюционного процесса.
В-четвертых, изучение генетики поведения животных важно для
отыскания новых методов наиболее рационального одомашнивания хозяй-
ственно полезных животных. Это имеет особенно большое практическое
значение для пушно-звероводческих хозяйств.
генетика поведения необходима для создания эксперимен-
тальных моделей нервных болезней. У мышей описано несколько десят-
ков неврологических наследственных болезней, которые изучаются в
качестве экспериментальных моделей болезней человека. Генотипичес-
ки обусловленная модель эпилепсии широко изучается на грызунах во
всех странах.
г. этому вопросу был посвящен интернациональный
коллоквиум во Франции.
Генетические исследования особенностей поведения начались вскоре
после вторичного открытия законов Менделя. Накопившийся к
щему времени материал показывает, что многие особенности поведения
наследуются по законам Менделя, однако в большинстве случаев ряд
факторов изменяет картину их наследования.
Для генетических исследований поведения удобной моделью оказа-
лись оборонительные реакции у животных. Этому вопросу посвящен ряд
исследований.
*Актуальные вопросы современной генетики. М.: Изд-во
1966. С.
174
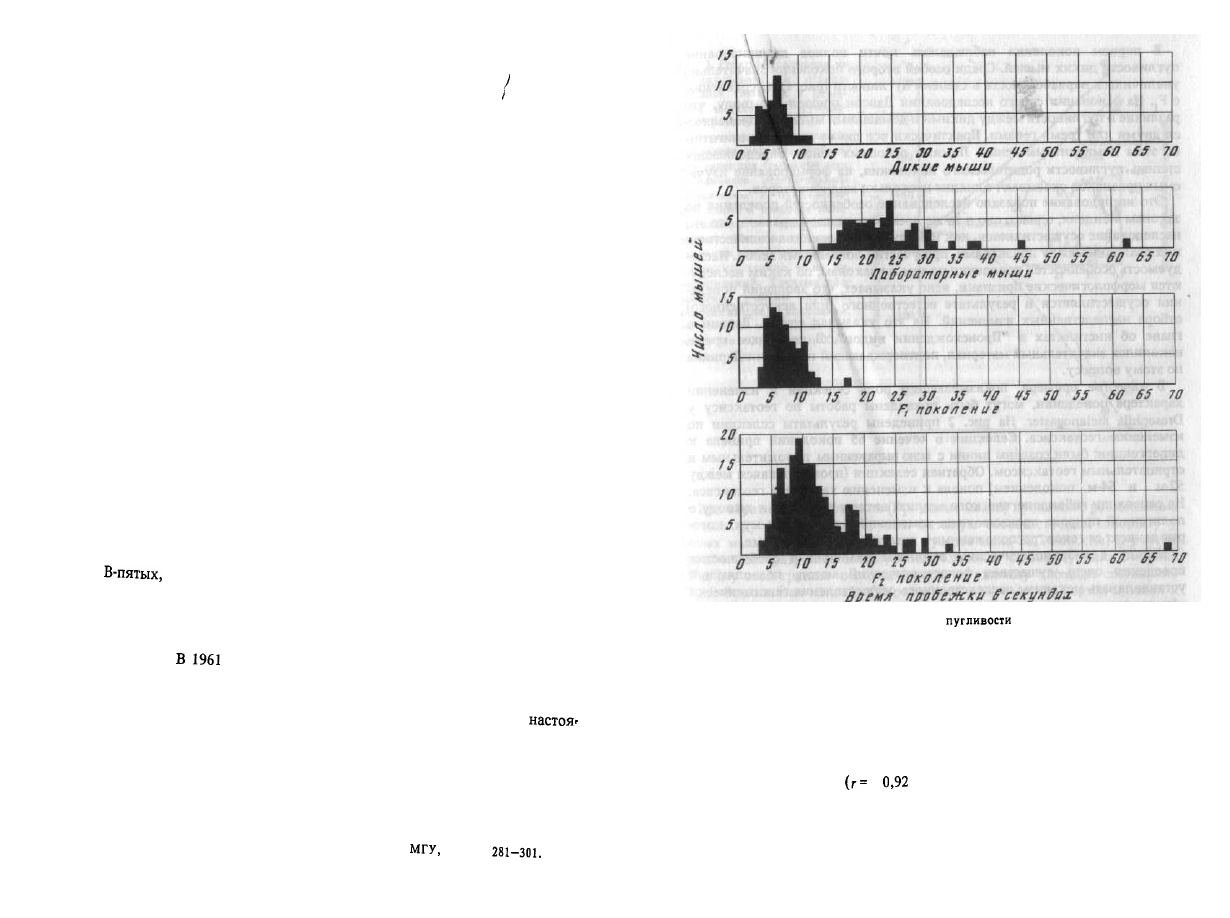
Рис. 1. Наследование
у мышей
В 1932 г. Давсоном были проведены исследования способа наследова-
ния резко выраженной пугливости диких мышей по сравнению со слабой
выраженностью этого признака у лабораторных мышей. Всего было
исследовано 3376 особей. Использовался объективный метод регистра-
ции: время пробежки по коридору (длиной 24 фута) при пугании мыши
передвигающимся движком. Предварительное исследование выявило
высокую корреляцию
+
± 0,003) между отдельными испытаниями
одних и тех же мышей, указывающую на значительную стабильность
изучаемых признаков поведения. Среднее время пробежки для диких
мышей составляло 5 с, для домашних - 20 с.
175