содержание .. 6 7 8 9 ..
Исследование по феногенетике признаков поведения у собак (В.П. Крушинский) - часть 8
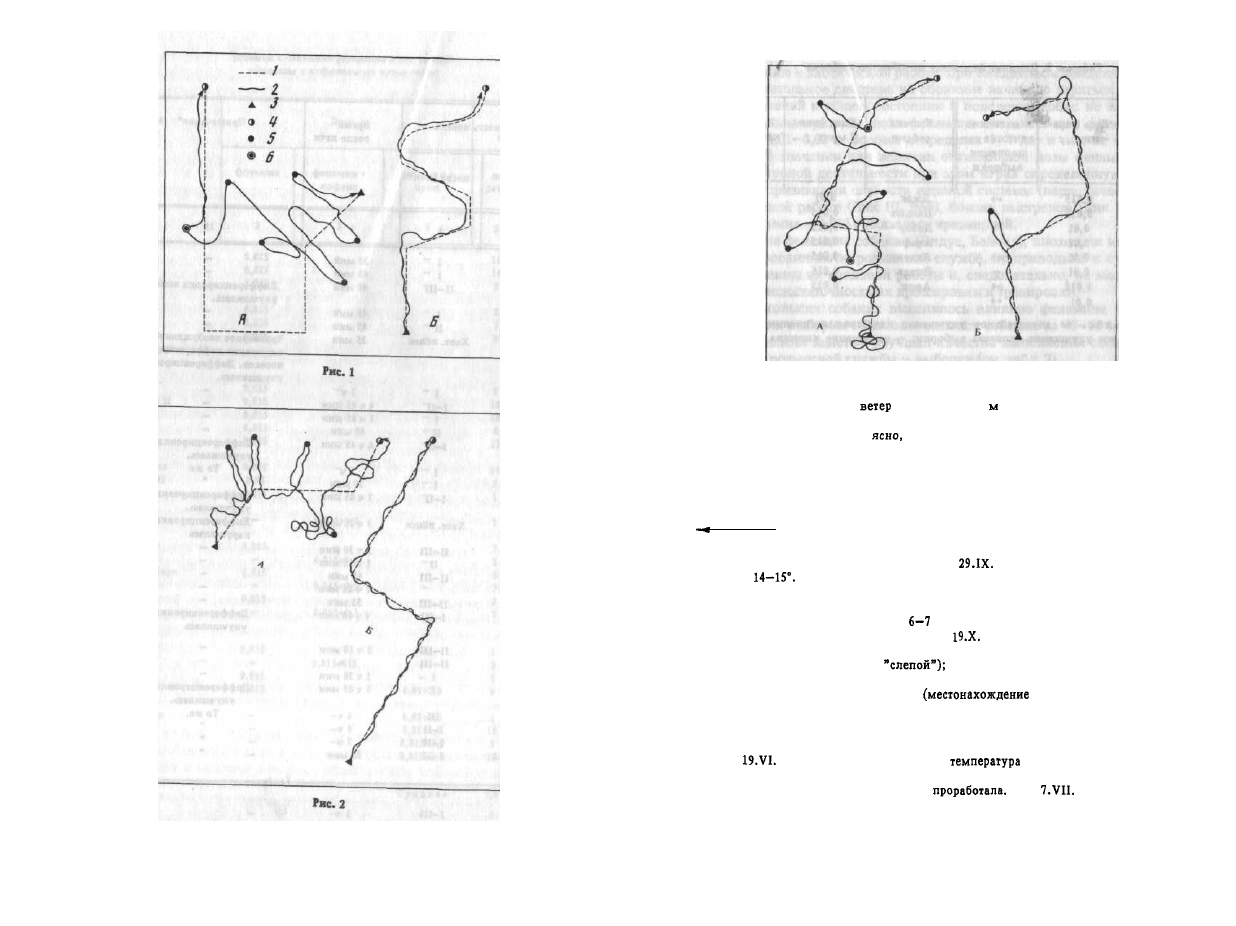
Рис. 3. Собака Верден
А — 8.VII. Погода: ясно,
слабый. 20 ч 55 проложен "слепой" след по траве
длиной 300 м; 21 ч 35 м пущена с исходной точки — собака практически почти не шла
по следу. Б — 8.VII. Погода:
ветер слабый. 22 ч 00 м. дан фенамин (0,015 г).
22 ч 37 м проложен "слепой" след длиной 300 м; 23 ч 12 м пущена с исходной точ-
ки — собака работала активнее и меньше отклонялась от следа, два угла проработала
самостоятельно
Рис. 1. Собака Дик
А — работа по следу до дачи фенамина.
Погода: ясно, ветер слабый, тем-
пература
16 ч Об м проложен след по травянистой поверхности длиной 500—
600 м (след "слепой" — направление такого следа проводнику неизвестно, и собака
должна прорабатывать его самостоятельно); 17 ч 07 м пущена с исходной точки —
вначале пошла правильно, но через
м сбилась и больше не могла отыскать след.
£ - работа по следу после дачи фенамина.
Погода: облачно, температура 7-8°.
Ветер слабый. 9 ч 55 м дан фенамин (0,01 г); 12 ч 55 м проложен след по травянистой
поверхности длиной 400 м (след
13 ч 55 м пущена с исходной точки — шла
по следу правильно на всем протяжении. 1 — линия следа; 2 — путь собаки; 3 — на-
чальная точка; 4 — конечная точка
помощника); 5 — место, с
которого контролирующий возвратил собаку на след; 6 — проводнику собаки указано
направление следа
Рис. 2. Собака Бой
А -
Погода: ясно, ветер слабый,
14-16°. 21 ч 40 м проложен
"слепой" след длиной 350 м по влажной траве; 22 ч 15 м пущена с исходной точки —
часто сбивалась и практически след не
£ —
Погода: ясно, ветер
слабый, температура 16°. 20 ч 30 м дан фенамин (0,015 г); 21 ч 15 м проложен "сле-
пой" след в поле по влажной траве длиной 600 м; 21 ч 50 м пущена с исходной точ-
ки - активно и правильно проработала след. Условные обозначения те же, что и
на рис.
1
119