содержание .. 1 2 3 4 ..
Исследование по феногенетике признаков поведения у собак (В.П. Крушинский) - часть 3
Следующий вопрос, который требовал выяснения: какова обучаемость
аппортировке у собак, обладающих стремлением носить предметы до
всякого
Три собаки с подобным поведением (Чанг, Альфа, Джульбарс) были
дрессированы аппортировке. Эти собаки чрезвычайно легко дрессирова-
лись этому приему. Две из них, Джульбарс и Чанг, с первого дня занятий
начали подносить предметы. Третья собака, Альфа, хватала предметы с
первого дня обучения, однако подносить их по команде начала со второ-
го занятия. Дальнейшим этапам обучения дрессировались только две
собаки (Альфа, Джульбарс). Обе эти собаки очень легко выдрессирова-
лись всем четырем этапам. Альфе понадобилось всего 10 занятий
(наименьшее число занятий из всех дрессированных собак). Джульбарс
выдрессировался на 15-й день занятий.
Таким образом, собаки, имеющие стремление носить предметы, обуча-
ются и самой аппортировке и всему, что с ней связано, чрезвычайно
легко.
По данным Шама, среди охотничьих собак имеется специально выве-
денная в Англии порода -
- для аппортировки убитой дичи.
Собаки этой породы - великолепные
обладающие,
помимо этого свойства, еще умением совершенно не мять подносимую
дичь, качеством, особенно ценимым охотниками у ретриверов.
Ретривер был выведен путем скрещивания некоторых пород охотни-
чьих собак и Лабрадора, породы собак, которая еще в начале XIX столе-
была ввезена в Англию и известна
как очень хороший
который, согласно Хагену, должен был передать эти
способности ретриверу. Данные о большой легкости, с которой ретривер
дрессируется аппортировке, мы находим у Дарвина.
Эти данные полностью согласуются с нашим выводом о наличии у
собак наследственно обусловленного различия в быстроте выработки
такого условного рефлекса, как подношение хозяину тех или других
предметов.
Может возникнуть вопрос: не является ли данное поведение собак,
обучающихся легко аппортировке и имеющих стремление носить предме-
ты, результатом общих свойств их нервной системы, связанных с повы-
шенной способностью к выработке условных рефлексов вообще, и не
является ли быстрая обучаемость аппортировке лишь одним из проявле-
ний этого свойства? Полученные мною данные говорят против такого
предположения.
Одна из упомянутых выше собак, Дже-Амур, обладавшая стремлением
носить предметы, была исследована мной в условно-рефлекторной
камере по обычной слюнно-секреторной методике.
Работа в камере показала, что Дже-Амур вырабатывал условные
рефлексы со средней скоростью. Так, например, первый условный реф-
лекс на метроном, 120 ударов в минуту (отставление 30
начал выраба-
тываться на
опыте
сочетаний) и достиг значительной прочности на
опыте (58-64
Второй условный рефлекс на звонок
щий) выработался после 20 сочетаний.
оказалась хорошей,
38
давая частые 0. По типу нервной деятельности собака может быть отнесена к
сангвиникам с довольно хорошим тонусом коры головного мозга. Никаких
особенных отклонений в
деятельности не было обнару-
жено.
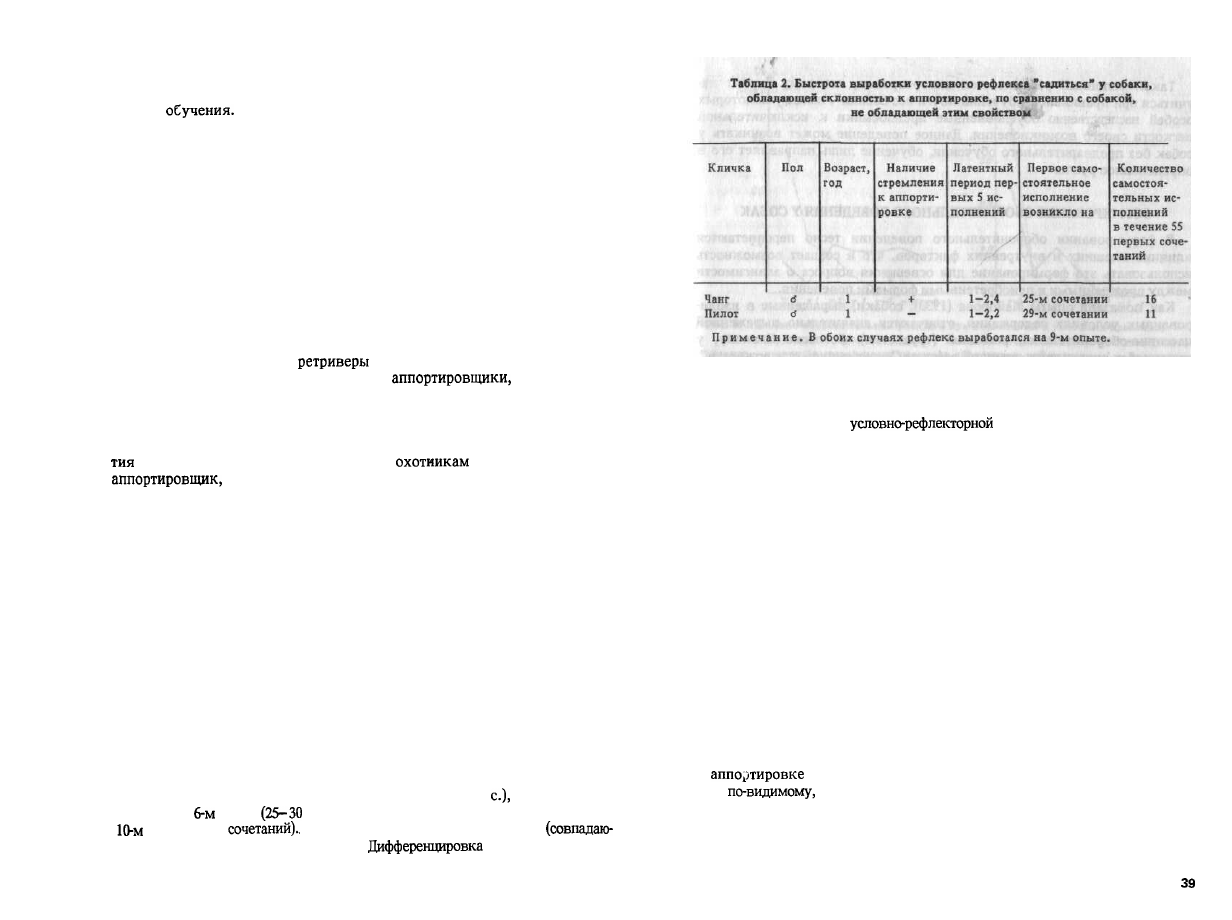
Вторая собака, Чанг, обладавшая стремлением носить предметы и
начавшая почти без всякого обучения аппортировать, была дрессирована
приему садится по команде "сидеть" с точной реакцией хода обучения. В
табл. 2 приведены данные хода обучения этой собаки по сравнению с
собакой, не обладающей стремлением носить предметы. Безусловный
раздражитель: нажим рукой на крестец и рывок за поводок.
Как видно из табл. 2, выработка двигательных условных рефлексов у
обеих собак происходила довольно сходно. Несколько более быстрый
ход выработки рефлекса у Чанга не настолько велик, чтобы его можно
было принять во внимание. Таким образом, у собак, обладающих стрем-
лением к аппортировке, вырабатываются условные рефлексы, не связан-
ные с ней, в пределах обычной нормы. Условные рефлексы, связанные с
аппортировкой, вырабатываются у них исключительно легко.
Итак, приведенные нами данные показывают следующее. У собак
имеется значительная изменчивость в быстроте и легкости выработки
двигательного условного рефлекса аппортировки предметов. В крайней
форме выражение этого свойства проявляется в стремлении собак без
предварительного специального обучения носить в зубах различные
предметы. Данное свойство обусловливается генотипически. Стремление
к
и легкость выработки условных рефлексов, связанных с
ней,
не являются выражением повышенной легкости и быстроты
выработки у таких собак всех рефлексов. Данное свойство является специфи-
ческой особенностью собаки, проявляющейся на фоне нервной деятельности, в
остальном обычной.