содержание .. 3 4 5 6 ..
Реакции и поведение собак в экстремальных условиях (М. Герд) - часть 5
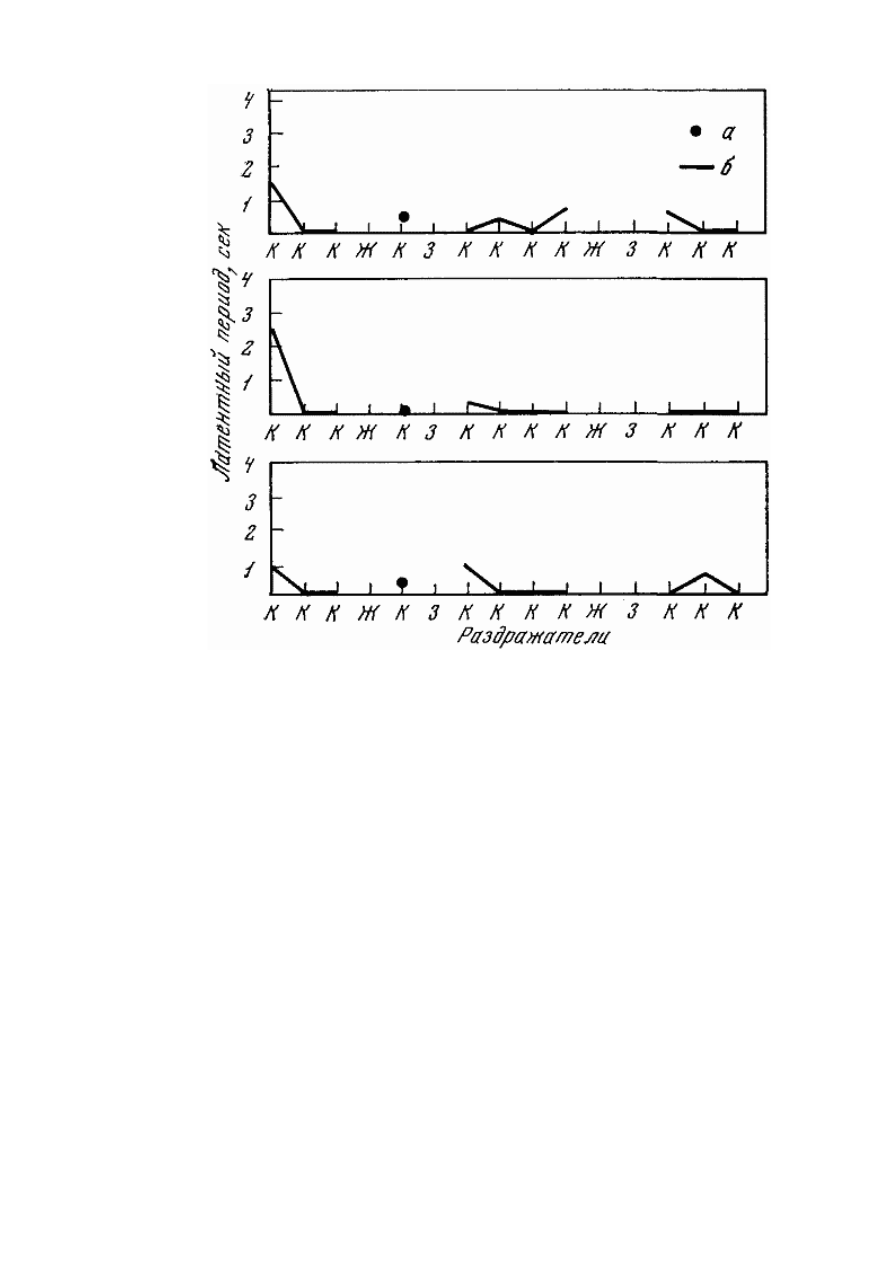
Рис. 22. Обследование высшей нервной деятельности путем трехкратной подачи
выработанного стереотипа раздражителей
Собака Стрелка (2. VI 1960 г.); а, б — рефлексы на положительные сигналы
В наших условиях использование существующих лабораторных стандартов для
определения типов было не применимо по ряду причин. Использование дрессировки лишало
возможности охарактеризовать процесс выработки условных рефлексов. Угашение с
подкреплением и без подкрепления и т. д. мы не могли применить, так как в каждый момент
необходимо было иметь хорошо закрепленные реакции на стереотип раздражителей и т. д. В
литературе (Паршутин и др., 1954; Флес, 1958) описывались приемы более простой и
доступной оценки типологических особенностей животных. Но эти методы также не могли
быть применены в наших условиях.
В связи со сказанным нам пришлось на основании принципов павловских
функциональных проб продумать свою систему проверки особенностей высшей нервной
деятельности собак. Оценочная роль такой системы не могла быть детально изучена.
Поэтому целью исследований было охарактеризовать, хотя бы частично, процессы
возбуждения, торможения и их баланса. Это позволяло сделать обоснованное заключение об
отсутствии у подопытных животных патологических черт реакций центральной нервной
системы.
Обследование нервной деятельности животных включало трехкратную подачу
выработанного стереотипа, пробу для определения силы возбудительного процесса, две
пробы для определения тормозного процесса и две — для испытания баланса
возбудительных и тормозных процессов.