Глава 5
Многослойный палеолитический комплекс в центральном зале Денисовой пещеры
5.1. Геоморфологическое положение и общая характеристика карстовой полости
В геоморфологическом отношении долина Ануя в окрестностях Денисовой пещеры характеризуется переходом от расширенного участка (Черноануйская депрессия) к суженному. Левый борт долины опирается на склоны горы Каракол (1 315м), правый борт -на склоны горы Сосновая (1 112м). Долина имеет асимметричный, близкий к V-образному поперечный профиль (рис. 28). Ширина днища долины около 120 м. Абсолютная отметка уреза воды 662 м. Левый борт долины слабовогнутый, крутизной в верхней части 25-30°, в нижней - до 15 - 20°. Правый борт долины выпуклый, крутизной в верхней части 30 - 35°, переходящий в нижней части в субвертикальные стенки высотой до 10 - 15 м. Правый борт рассечен короткими сухими распадками и осыпными лотками, заложившимися по зонам трещиноватости коренных пород. Сливающиеся конусы выноса и осыпи отжимают современное русло Ануя к противоположному борту и формируют единую аккумулятивную поверхность протяженностью 80 - 100 м с уклоном 15 - 20° к руслу. Склоны по обоим бортам покрыты лесом: по левому борту - березово-лиственничным, по правому - разреженным березово-сосновым с кустарником в подлеске.

Рис. 28. Вид на долину Ануя из Денисовой пещеры.

Рис. 29. Вид на Денисову пещеру
Пещера выработана в правом борту долины Ануя в крупном блоке силурийских биогермных известняков. Вход в пещеру расположен в уступе отвесной стены юго-западной экспозиции на высоте около 30 м над современным урезом реки (рис. 29). Пещера состоит из системы коротких субгоризонтальных и пологона-клонных галерей различных размеров, сообщающихся через центральный зал (рис. 30). До начала раскопок вход в пещеру представлял овальное отверстие размером 7 х 1,7 м. В процессе раскопок и удаления части рыхлых отложений высота входа увеличилась и достигает в настоящее время 6 м. Вход ведет в главную галерею шириной до 7 м и протяженностью в северо-западном направлении около 10м, которая под прямым углом открывается в центральный зал пещеры. Из центрального зала одна галерея длиной 9 м и шириной от 1 до 4,5 м идет в юго-западном направлении, открываясь на предвходовую площадку. Две узкие темные галереи уходят в юго-восточном направлении в глубь карстового массива на 50 - 70 м, где полностью перекрыты рыхлыми отложениями. Общая площадь пещеры составляет 270 м2, объем - 330 м3. При этом суммарная площадь центрального зала и главной галереи равна 120м2.
Центральный зал представляет сводовую камеру с размерами в плане 11 - 9 м и высотой около 10 м. В южной части свода центрального зала находится сквозное отверстие около 1 м в поперечнике. Стены пещеры выровненные, заглаженные. На них хорошо читается сложный рисунок системы пологонаклонных и субвертикальных трещин, определивших, вероятно, структурный план пещеры. Большинство трещин залечено карбонатным цементом. Натечные образования в центральном зале и подводящих галереях отсутствуют, лишь под слоем рыхлого гумусированного материала, покрывающего днище пещеры, сохранились участки развития тонких травертиновых корочек и коррозионных желобков.
В настоящее время образования в коренных породах новых трещин или расширения старых не происходит. Об этом можно судить, внимательно исследовав проведенную в 1984 г. масляной краской по обеим стенам главной галереи условную "нулевую" линию, служащую своеобразным репером взаимного расположения отдельных блоков. Пересекающая многочисленные трещины полоса нигде не разорвана и не смещена, а слой краски, заполнивший открытые трещины, не поврежден.
За нулевую линию принята отметка кровли голоценовых отложений у наиболее отдаленной от капельной линии северо-восточной стены центрального зала пещеры (рис. 31). Превышение нулевой линии над урезом Ануя по данным инструментальной съемки составляет 31,7м. Глубина залегания слоев в стратиграфических разрезах указана относительно нулевой линии пещеры.
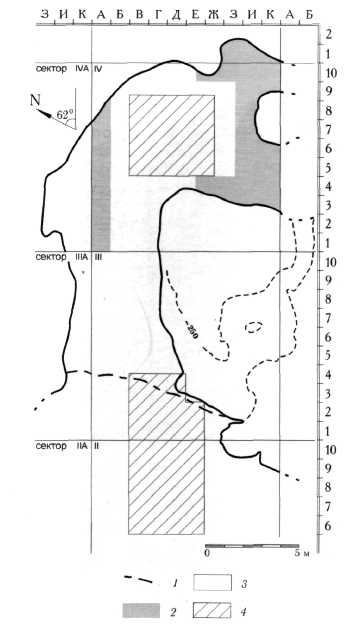
Рис. 30. План Денисовой пещеры. 1 - капельная линия, 2 - современная поверхность, 3 - площадь вскрытых голоценовых отложений, 4 - площадь вскрытых плейстоценовых отложений.

Рис. 31. Продольный разрез полости Денисовой пещеры.
5.4. Фауна крупных млекопитающих палеолита Северо-Западного Алтая
Костные остатки крупных и средних млекопитающих принадлежат 27 видам из отрядов: Carnivora -Alopex lagopus, Vulpes corsac, Vulpes vulpes, Cuon alpinus, Canis lupus, Ursus arctos, Ursus rossicus, Martes zibellina, Mustela eversmannii, Mustela erminea, Mustela altaica, Mustela nivalis, Crocuta spelaea, Panthera spelaea, Lynx lynx; Proboscidea - Mammuthus primigenius; Perissodactyla - Coelodonta antiquitatis, Equus hydruntinus, Equus ferus; Artiodactyla -Capreolus pygargus, Cervus elaphus, Poephagus mutus, Bison priscus, Procapra gutturosa, Saiga tatarica, Capra sibirica, Ovis ammon.
Среди фаунистических материалов по числу видов (исключая эврибионтные) и количеству костных остатков преобладают обитатели степных пространств (Vulpes corsac, Mustela eversmannii, Ursus rossicus, Crocuta spelaea, Coelodonta antiquitatis, Equus hydruntinus, Poephagus mutus, Bison priscus, Procapra gutturosa, Saiga tatarica, Ovis ammon). Вместе с ними найдены останки представителей лесостепных (Сиоп alpinus, Equus ferus, Cervus elaphus) и скальных (Mustela altaica, Capra sibirica) видов. Останки обитателей леса (Martes zibellina, Ursus arctos, Lynx lynx, Capreolus pygargus) присутствуют практически по всему разрезу, за исключением слоя 9. Кроме того, в слоях 22, 19, 12-9 обнаружены единичные останки представителей тундрово-таежных видов (Alopex lagopus, Mustela erminea). В целом видовой состав териофауны свидетельствует о мозаичности ландшафта, характерной для горного рельефа. Принципиальных изменений в видовом составе крупных млекопитающих на разных стратиграфических горизонтах не наблюдается, хотя вверх по разрезу можно отметить определенные различия в численности костных остатков у видов разных биотопических групп.
В экологическом спектре слоя 22 присутствуют все группы животных, при этом кости степных видов составляют 58,8%. Следующую по значению группу образуют лесные виды, на долю которых приходится 13,7% костных остатков. Обитателям скал принадлежит 11,8% определимых костей. Виды лесостепи и тун-дрово-таежной группы представлены небольшим количеством останков. На их долю приходится по 7,8% костей. Характерной чертой тафономии этого слоя является заметное преобладание костных останков медведей: они составляют 48,8% всех определимых костей в слое и 71% всех медвежьих останков, обнаруженных в пещере.
Распределение костного материала в отложениях средней части разреза отражает общее господство млекопитающих открытых пространств и неуклонную тенденцию сокращения снизу вверх численности лесных форм. Фаунистические материалы слоев 21 и 20 относительно малочисленны. При их суммарной характеристике наибольший процент (78,9) составляют останки представителей степных форм; на лесные виды приходится 10,5%; обитателям скал и лесостепи принадлежит по 5,3% костей. Выше по разрезу, в наиболее насыщенных костями слоях 19, 14, 12 и 11 отмечены следующие соотношения остатков костей животных различных экологических групп: 1) степные-слой 19-72,7%, слой 14-67,5%, слой 12-73,3%, слой 11-71,4%; 2) лесостепные - слой 19-10,2%, слой 14-10,4%, слой 12-13,3%, слой 11-6,5%;3)скальные-слой 19-8%, слой 14 - 9,1 %, слой 12 - 9,5%, слой 11-17,5%; 4) лесные - слой 19 - 6,8%, слой 14 - 13%, слой 12 - 1,9%, слой 11 - 3,9%; 5) тундрово-таежные - слой 19 - 2,3%, слой 14- 0%, слой 12 -1,9%, слой 11 - 0,6%. Изменение количественного показателя останков Crocuta spelaea и Equus hydruntinus как более теплолюбивых видов в составе обитателей степных ландшафтов (слои 21,20-43,3%, слой 19-51,6%, слой 14-32,7%, слой 12-27,3%, слой 11 - 28,2%) предполагает относительное ухудшение климатических условий вверх по разрезу.
В верхней части плейстоценовой толщи (слой 9) на долю животных степных и лесостепных биотопов приходится соответственно 64,3 и 7,1% костей. Удельный вес ископаемых останков тундрово-таежных форм составляет 2,4%. Лесные виды в этом слое отсутствуют, а количество костей обитателей скал заметно увеличилось (26,2%), что, возможно, было вызвано снижением границ высотных поясов. Вместе с тем здесь по сравнению с нижележащими слоями 12 и 11 показатель более теплолюбивых видов среди степных зверей (22,2%) существенно не меняется.
Детальная характеристика костных остатков крупных млекопитающих из плейстоценовых отложений Денисовой пещеры представлена в главе 7.
5.5. Фауна мелких млекопитающих палеолита Северо-Западного Алтая
Для изучения плейстоценового сообщества мелких млекопитающих использованы фаунистические материалы двух участков:
- раскоп 1994 г. - квадраты Е/6-8 (линия субквадратов а - в), слои 9-22 общей мощностью 4 м;
- раскоп 1995 г. - квадраты Е/5 (субквадраты а - г), Е/6-8 (линия субквадратов б - г), слои 9-22 общей мощностью 4,5 м.
Результаты определения собранных материалов и заключения по ним приведены для каждой коллекции отдельно, что обусловлено рядом причин. Отложения Денисовой пещеры отличаются сложным пространственным залеганием. Ряд слоев имеет линзовидный характер, а некоторые слои перемяты и смещены за счет конвективной неустойчивости грунтов. От квадрата к квадрату меняется мощность слоев, нарушается их последовательность за счет выклинивания некоторых горизонтов. Также неравномерна концентрация костного материала по площади. Эти обстоятельства придают высокую тафономическую индивидуальность отдельным участкам раскопа. Фактически каждый квадрат неповторим по условиям осадконакопления и набору костных остатков млекопитающих. Учитывая сложную тафономическую ситуацию, необходима дифференцированная оценка полученных результатов. Такой подход позволяет отделить случайные флуктуации таксономического разнообразия от истинных изменений состава фауны, отражающих динамику развития природной среды.
Общий список таксонов мелких позвоночных, установленный для плейстоценовых отложений Денисовой пещеры, включает более 40 названий (табл. 3).
В коллекции 1994 г. количество определимых костных остатков достигает 7 тыс. экз. При оценке относительного обилия различных таксонов было использовано 6 326 экз. В среднем на каждый слой приходится приблизительно 200 определимых остатков. В целом нижняя часть разреза охарактеризована более надежно, чем верхняя. Характер распределения костных остатков по стратиграфическим подразделениям разреза представлен на диаграммах общего (рис. 46) и выборочного (рис. 47) состава мелких животных. За время накопления толщи плейстоценовых отложений видовой состав млекопитающих менялся незначительно. Этот процесс отражает сходный механизм формирования танатоценоза пещеры и сходный состав населения мелких млекопитающих в долине Ануя в период накопления толщи слоев 22-9.
На фоне относительной стабильности состава фауны от нижних слоев к верхним обращает на себя внимание резкое уменьшение количества останков летучих мышей Chiroptera в отложениях слоя 21. Выше слоя 19 их количество сокращается приблизительно в десять раз и на этом уровне сохраняется до кровли плейстоценовых отложений. Объяснение этого феномена с палеоэкологических позиций найти трудно. Судя по составу мелких млекопитающих, от слоя 22 вверх по разрезу внешние природные факторы не претерпели существенных изменений. Видимо, единственной причиной, нарушившей условия обитания летучих мышей в пещере, был человек. Можно предполагать, что, начиная с эпохи накопления слоя 21, присутствие человека в пещере стало постоянным, а использование огня -регулярным. Это предположение подтверждается та-фономическими наблюдениями. Первые хорошо выраженные прослойки углей появляются в отложениях слоя 21. Начиная со слоя 20, резко возрастает количество артефактов палеолитического человека. Эти данные свидетельствуют о стабильном присутствии человека и задымлении полости пещеры, что привело к увеличению фактора беспокойства и к резкому сокращению численности летучих мышей в пещере.
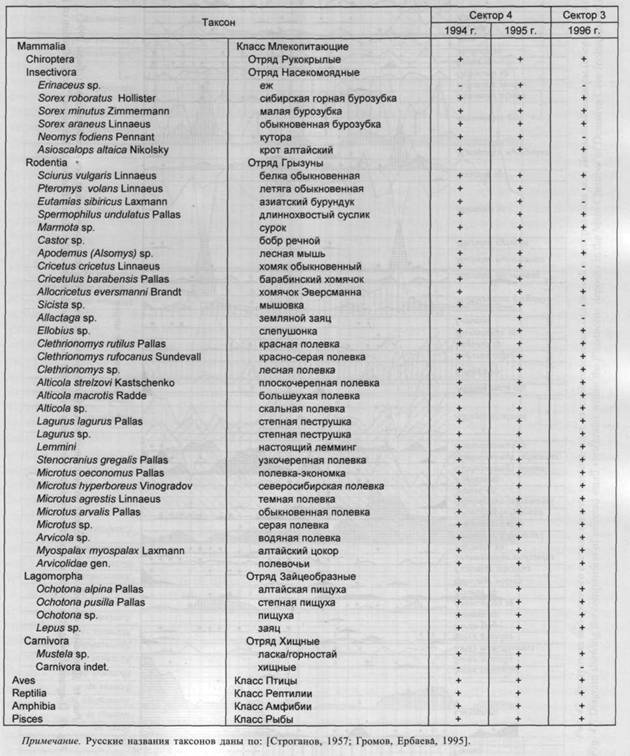
Таблица 3. Общий состав мелких позвоночных из плейстоценовых отложений в Денисовой пещере

Рис. 46. Диаграмма общего состава мелких позвоночных из плейстоценовых отложений в центральном зале Денисовой пещеры (сборы 1994 г.).
 Диаграмма
выборочного состава мелких позвоночных из плейстоценовых отложений в центральном
зале Денисовой пещеры
Диаграмма
выборочного состава мелких позвоночных из плейстоценовых отложений в центральном
зале Денисовой пещеры
Остальная часть микротериофауны дает ясную картину единого сообщества, однако его структура постепенно менялась во времени. Причем состав этого сообщества принципиально отличался от современного, что свидетельствует об ином облике окружающих ландшафтов в плейстоцене.
Ведущую группу мелких млекопитающих на протяжении палеолитического периода составляли полевки Microtinae. В питании хищных птиц наибольшую численность имели азиатские горные полевки рода Alticola, среди последних доминировала плоскочерепная полевка Alticola strelzovi, обитатель каменистых сухих склонов, покрытых редкой растительностью. Количественное соотношение этой полевки с другими видами достаточно стабильно и варьирует по горизонтам от 10 до 20%, несколько увеличиваясь в слоях 11-9 до 29%. Содоминантами в составе фауны являлись узкочерепная полевка Stenocranius gregalis и рыжие полевки Clethrionomys. В целом численность узкочерепной полевки несколько выше численности рыжей полевки. По-разному выглядит тенденция их динамики во времени. Численность узкочерепной полевки вверх по разрезу постепенно нарастает, а рыжей полевки -снижается. Среди останков рыжих полевок преобладают зубы Clethrionomys rutilus, изредка встречаются зубы С. rufocanus.
В плейстоценовых отложениях присутствуют степные пеструшки рода Lagurus. Их останки впервые появляются на уровне 10 горизонта 22.2. Однако до слоя 21 обилие пеструшек не превышает 1% от общего количества останков. Начиная со слоя 20, их численность постепенно нарастает, давая максимум в слое 9-11% от общего числа останков. По своей морфологии ископаемые пеструшки из неоплейстоценовых отложений пещеры близки современной Lagurus lagurus.
Останки полевки-экономки Microtus oeconomus составляют также незначительную долю в выборке -от 0,5 до 3%. Однако они встречаются стабильно, практически во всех горизонтах. Постоянно отмечаются землеройки рода Sorex, хотя обилие их останков не превышает 2-3%. Сходную картину дают суслики Spermophilus undulatus, даурский хомячок Cricetulus barabensis и хомяк Эверсманна Allocricetus eversmanni, останки которых отмечены во всех горизонтах. Однако ни те ни другие не найдены на уровне 13 горизонта 22.3. Кроме того, встречаемость суслика нарастает вверх по разрезу, достигая максимума в горизонте 9.1. Численность даурского хомячка стабильна в средней части разреза. Хомяк Эверсманна дает пик в горизонте 9.1.
Другая группа мелких млекопитающих, останки которых постоянно встречаются в плейстоценовых отложениях Денисовой пещеры, - зайцеобразные. Среди них ведущее место занимает пищуха Ochotona. Ее останки найдены практически во всех горизонтах и составляют от 1 до 6%. Причем наибольшее количество приходится на нижнюю часть горизонта 22.1 (уровни 3 - 5). Всплеск численности фиксируется также в горизонтах 11.2 и 9.2. Полученный материал лишь в некоторых случаях позволяет диагностировать виды. Отмечено присутствие двух групп: мелкой степной пищухи Ochotona pus ilia и более крупной О. alpina. Однако количество определимых до вида останков недостаточно, чтобы анализировать различия в их послойной приуроченности.
Регулярно в отложениях пещеры встречаются останки зайцев. Их относительное количество колеблется в пределах 0,15 - 3%. В небольшом количестве, но по всей толще пещерных отложений, встречаются останки мелких хищников, преимущественно куньих. Их основная часть принадлежит представителям рода Mustela.
Третью группу в списке мелких млекопитающих составляют виды, количество костных остатков которых относительно невелико, а встречаемость по слоям спорадична. К ним относится сурок Marmota, число останков которого колеблется в интервале 0,2 - 1,2%. Останки сурка зарегистрированы на уровнях 10, 8 - 5, 1 слоя 22, в слое 20 и в горизонтах 19.1,12.1 и 9.3. Их небольшое количество и плохая сохранность не позволяют диагностировать вид. Вероятно, в танатоценоз пещеры сурок попадал не в погадках хищных птиц, а с пометом млекопитающих (волка, гиены) или как охотничий трофей человека. По нашим наблюдениям, сурок был объектом охоты палеолитического человека на стоянке Каменная Балка в Приазовье. Известно, что сурок добывался охотниками некоторых палеолитических стоянок Франции [Patou, 1987]. Еще реже и в меньшем количестве встречается бурундук Eutamias sibiricus. Он отмечен на уровнях 12,6,4 и 3 слоя 22 и в горизонте 19.1. В двух случаях отмечены белка Sciurus (уровни 6 и 4 слоя 22) и летяга Pteromys volans (уровень 12 горизонта 22.3 и горизонт 13.1).
В общей динамике численности мелких млекопитающих некоторое своеобразие имеет слепушонка Ellobius. Впервые ее останки появляются в горизонте 19.1. Далее по разрезу они отмечены в горизонтах 13.1,12.3,9.3 и 9.1. В горизонте 12.3 их количество достигает 2,4%.
Редкими представителями тафоценоза плейстоценовых слоев пещеры являются серый лемминг, темная полевка и водяная полевка. Представители родаЬеттиз найдены на уровнях 12, 10, 9 слоя 22 и в горизонтах 13.4,13.1,12.3. Число их останков колеблется от 0,2 до 0,95%. В четырех случаях найдена темная полевка Microtus agrestis - уровни 12,11 горизонта 22.3, горизонты 19.1 и 13.4. Водяная полевка Arvicola, близкая по своим морфологическим признакам современной Arvicola terrestris, отмечена в слоях 20,19,17, а также в горизонтах 13.3. и 9.2. К наиболее редким представителям ископаемой фауны принадлежат копытный лемминг и лесная мышь. По одному фрагменту зубов копытного лемминга Dicrostonyx отмечено в горизонтах 19.1 и 12.1. Мыши рода Apodemus найдены в нижней части слоя 22 (уровни 11 и 10).
Помимо мелких млекопитающих в формировании состава тафоценоза палеолитических слоев принимали участие останки птиц, рыб, отчасти лягушек и мелких рептилий. Число останков птиц варьирует по разным слоям от 0,5 до 9,3%. Позвонки и отдельные кости рыб найдены во всех горизонтах разреза. Их относительное количество колеблется от 6,5 до 23%, составляя в среднем 7 - 9%. Останки лягушек и рептилий встречаются в небольшом количестве и спорадично по отдельным горизонтам разреза.
В коллекции 1995 г. общий объем определимых костных остатков составил 27 663 экз. Наиболее репрезентативный материал получен из отложений горизонта 22.1, а также в интервале горизонтов 12.1-11.1. Характер распределения костных остатков по стратиграфическим уровням разреза 1995 г. представлен на диаграммах общего (рис. 48) и выборочного (рис. 49) состава мелких позвоночных. Видовой состав мало меняется вверх по разрезу. Единственная группа мелких млекопитающих, количество останков которой резко меняется на границе слоев 22 и 21, - летучие мыши. Остальные виды позвоночных не претерпели резких колебаний численности в период накопления всей толщи. В целом ископаемая фауна Денисовой пещеры принципиально отличается от современной. Она отражает значительно большее разнообразие биотопических условий и пестроту ландшафтов по сравнению с современностью. Это разнообразие было обусловлено заметным участием в сообществах степных и высокогорных элементов.
 Рис.
48. Диаграмма общего состава
мелких позвоночных из плейстоценовых отложений в центральном зале Денисовой
пещеры (сборы 1995 г.).
Рис.
48. Диаграмма общего состава
мелких позвоночных из плейстоценовых отложений в центральном зале Денисовой
пещеры (сборы 1995 г.).
 Рис.
49. Диаграмма выборочного состава
мелких позвоночных из плейстоценовых отложений в центральном зале Денисовой
пещеры (сборы 1995 г.). 1 -
деформация, 2 - содержание менее 1%.
Рис.
49. Диаграмма выборочного состава
мелких позвоночных из плейстоценовых отложений в центральном зале Денисовой
пещеры (сборы 1995 г.). 1 -
деформация, 2 - содержание менее 1%.
Несмотря на общую однородность сообществ мелких млекопитающих, благодаря большому объему материала, удалось наметить определенную динамику количественного соотношения отдельных таксонов. Анализ этих флюктуации позволил проследить общие закономерности изменения биоты за время накопления плейстоценовых осадков Денисовой пещеры.
Летучие мыши Chiroptera являются единственным компонентом тафоценоза, который резко меняет свою численность. В слое 22 их относительное количество колеблется от 14,5% на уровнеЗ до 63,8% на уровне 13. К слою 21 обилие летучих мышей снижается до 9,9%. Далее по разрезу оно варьирует в интервале 3 - 6,6% до слоя 17, а в верхней части разреза их численность составляет менее 3%. Эти данные подтверждают отмеченную ранее закономерность и придают ей более достоверный характер. Скорее всего, на позднем этапе накопления осадков слоя 22 произошло принципиальное изменение в характере присутствия в пещере первобытного человека. Менее понятны два минимума численности летучих мышей на уровнях 8 и 3 слоя 22. При этом падение численности от уровня 13 к уровню 8 и от уровня 6 к уровню 3 имеет закономерный характер, что должно быть связано с природными явлениями, а не с чисто тафономическими причинами. Падение численности летучих мышей на уровне 8 слоя 22 отмечено также в материалах 1994 г., что существенно повышает уровень их достоверности.
Все летучие мыши из плейстоценовых отложений Денисовой пещеры являются видами, которые используют пещеры в качестве убежища. Видимо, накопление их костных остатков происходило в результате естественной гибели зверьков из состава зимующих колоний. Небольшая часть материала имеет следы обработки желудочным соком и, следовательно, попала в состав тафоценоза из погадок хищных птиц. В ископаемой фауне летучих мышей Денисовой пещеры установлено семь видов: остроухая ночница Myotis aff. blythi Tomes, прудовая ночница М. dasycneme Boie, ночница Брандта М. aff. brandti Eversmann, водяная ночница М cf. daubentoni Kuhli, сибирский (большой) трубконос Murina leusogaster Milne-Edwards, бурый ушан Plecotus auritus Linnaeus, северный кожанок Eptesicus cf. nilssoni Keyserling, Blasius.
Максимальное число определимых останков Chiroptera приходится на слой 22. В этом же слое наблюдается максимальное видовое разнообразие летучих мышей. Здесь найдены останки всех рукокрылых, определенных для Денисовой пещеры. В отложениях горизонта 22.3 не найдена только Myotis cf. daubentoni, останки которой обнаружены в горизонте 22.1. Максимальное видовое разнообразие рукокрылых отмечено на уровне 6 горизонта 22.2. Здесь представлены пять видов летучих мышей; отсутствуют только остроухая ночница М. aff. blythi и водяная ночница М. cf. daubentoni. Наименьшее число видов отмечено на уровне 8 горизонта 22.2: Plecotus aff. auritus к Eptesicus cf. nilssoni. На этот же уровень приходится минимальная абсолютная численность останков Chiroptera. Наиболее многочисленны в слое 22 останки ушана Plecotus aff. auritus - около 60% от всего определенного материала по этому слою. Eptesicus cf. Nilssoni и Myotis aff. dasycneme принадлежит по 12% останков. Костные остатки Murina aff. leucogaster и Myotis aff. brandti составляют 3,5% . На долю Myotis aff. blythi ji M. cf. daubentoni приходится около 1% определенного по этому слою материала.
На рубеже слоев 22 и 21 резко падает абсолютное количество и видовое разнообразие останков рукокрылых. В слое 21 обнаружено только два вида летучих мышей - бурый ушан (Plecotus aff. auritus) и северный кожанок (Eptesicus cf. nilssoni). В отложениях слоев 19 и 17 отмечено небольшое количество останков Plecotus aff. auritus, Myotis blythi, Plecotus aff. auritus и М. aff. dasycneme. Верхняя пачка плейстоценовой толщи (слои 13-9) также бедна ископаемыми рукокрылыми. Здесь найдены останки четырех видов: Myotis aff. blythi, M. aff. dasycneme, Eptesicus cf. nilssoni и Plecotus aff. auritus.
Следует отметить, что для значительной части ископаемых форм летучих мышей Денисовой пещеры использовалась открытая номенклатура. Это связано с тем, что по своей морфологии они не полностью идентичны аналогичным современным видам. Например, зубы плейстоценового северного кожанка Eptesicus cf. nilssoni и ночницы Брандта Myotis aff. brandti значительно крупнее близких им современных форм.
Большинство ископаемых летучих мышей Денисовой пещеры в своей экологии связаны с широколиственными ил» хвойно-широколиственными лесами. Например, ночница Брандта населяет смешанные и широколиственные леса, по поймам проникает в тайгу и степь. Она распространена от Англии и востока Испании через Урал и Южную Сибирь до Кореи и Японии. Сибирский трубконос - обитатель смешанных хврйно-широколиственных и широколиственных (преимущественно горных) лесов от Алтая до Гималаев, Юго-Восточного Китая и Дальнего Востока. Северный кожанок рпсмроетраттеп п лесттой части Сеперпой Евразии: от Франции, Швейцарии и Кавказа до Гималаев, Тибета, Монголии и Дальнего Востока. На севере его ареал совпадает с границей лесной зоны. Водяная ночница является обитателем речных пойм лесной и лесостепной зон Европы, Южной Сибири; ее ареал простирается до Китая и Японии. Лишь остроухая ночница заселяет не только лесные, но и аридные ландшафты от Средиземноморья, через Кавказ, до Центральной Азии.
Еж Erinaceus sp. отмечен в плейстоценовой толще только в отложениях горизонта 14.2.
Землеройки Sorex sp. обычны в плейстоценовых отложениях пещеры, но ни в одном слое их количество не превышает 3%. Небольшое увеличение численности этих насекомоядных отмечено на уровнях 10, 8, 6 горизонта 22.2 и в слое 12. Подавляющая часть останков землероек принадлежит Sorex araneus. Изредка встречается мелкая S. minutus, еще реже - крупная 5. roboratus. Увеличение видового разнообразия землероек отмечено в отложениях слоя 12.
Кутора Neomys sp. очень редка в отложениях пещеры. Ее единичные фрагменты найдены в горизонтах 14.2 и 12.3. Скорее всего, это связано с тем, что кутора обитает в пойменных, хорошо защищенных биотопах.
Алтайский крот Asioscalops sp. найден практически во всех горизонтах плейстоценовой толщи. Максимальное число его останков приходится на слой 20-5,2%.
Белка Sciurus sp. очень редкий вид в отложениях пещеры. Ее единичные находки отмечены на уровне 5 горизонта 22.1 и в осадках горизонта 11.3.
Летяга Pteromys sp. также редкий вид. Ее останки обнаружены на уровнях 8,6 и 5 слоя 22, а также в отложениях горизонтов 19.1,14.2и 11.1.
Бурундук Eutamias sibiricus отмечен только на уровне 8 горизонта 22.2 и в отложениях слоя 21.
Длиннохвостый суслик Spermophilus undulatus обычен для всей толщи плейстоценовых отложений. Однако его относительная численность нигде не превышает 3%. Вероятно, это связано прежде всего с механизмом формирования тафоценоза: как объект охоты для птиц, гнездящихся в пещере, суслик был слишком велик.
Сурок Marmota sp. встречается с редкими перерывами почти по всему разрезу, за исключением нижней части слоя 22 (уровни 13-8).
Бобр Castor sp. известен только по сколам щечных зубов и эмали резцов, которые зарегистрированы в отложениях горизонтов 21.2 и 14.3.
Лесные мыши Apodemus (Alsomys) sp. определены по отдельным находкам в отложениях горизонтов 19.2. и 9.3
Хомяк обыкновенный Cricetus cricetus отмечен единичными останками на уровнях 5, 4, 1 горизонта 22.1 и в отложениях горизонтов 19.3,19.1. Эти материалы существенно дополняют сборы 1994 г., в которых хомяк был обнаружен только на уровне 3 горизонта 22.1.
Барабинский хомячок Cricetulus barabensis встречается по всей толще плейстоценовых отложений, хотя его относительная численность нигде не превышает 2 - 2,5%. В целом в нижних слоях он встречается чаще, а его количество несколько выше, чем в верхней части разреза.
Хомячок Эверсманна Allocricetus eversmanni также встречается почти во всех слоях разреза, но его количество не превышает 1,5- 2%. Этот вид, в противоположность барабинскому хомячку, проявляет более устойчивую численность п верхней части разреза.
Мышовка Sicista sp. является относительно редким видом. Ее останки найдены на уровне 7 горизонта 22.2, а также в отложениях горизонтов 21.1,12.3,12.1, 11.3 и 9.2. Численность мышовки колеблется в интервале 0,25 - 0,8%. В сборах 1994 г. этот грызун не зарегистрирован, что, возможно, объясняется чисто вероятностным фактором.
Тушканчик Allactaga sp. из группы земляных зайцев отмечен единичными зубами на уровнях 3 и 2 горизонта 22.1 и в отложениях горизонта 19.2.
Слепушонка Ellobius sp. спорадично встречается в средней и верхней частях разреза, начиная с уровня 1 горизонта 22.1.
Рыжие или лесные полевки Clethrionomys sp. составляют важнейший компонент тафоценозов Денисовой пещеры. Их костные остатки присутствуют во всех горизонтах разреза. Относительная численность рыжих полевок колеблется от 2 до 11%. Если учесть
результаты анализа современных погадок, то можно предполагать, что роль этих полевок в плейстоценовых сообществах долины Ануя была еще выше. В ископаемом состоянии, в отличие от современности, рыжие полевки представлены двумя видами - Clethrionomys rutilus и С. rufocanus.
Азиатские горные полевки AIticola sp. являются одной из самых массовых групп мелких млекопитающих плейстоцена. В среднем их численность составляет 10 -15% от общего количества мелких позвоночных, а в горизонтах 14.2 и 9.3 она превышает 20%. В ископаемой фауне азиатские горные полевки представлены двумя видами - Alticola strelzovi и A. macrotis. В современной фауне бассейна Ануя присутствует только первый вид -г плоскочерепная полевка.
Степная пеструшка Lagurus sp. - один из важных компонентов тафоценоза пещерных осадков. Этот вид встречается почти на всех горизонтах плейстоценовой толщи. Однако заметную роль пеструшка начинает играть в отложениях слоя 20. Здесь и выше по разрезу ее численность колеблется от 3 до 5%. В отложениях горизонта 9.2 количество пеструшки возрастает до 8,7%, а в осадках горизонта 9.1 достигает 17,9%. Важно отметить, что подобная тенденция отмечена в динамике численности по сборам 1994 г. Сходство проявляется и в заметном падении численности в интервале горизонтов 13.4- 12.3. Это сходство придает особую достоверность полученным результатам. В целом ископаемые пеструшки представлены видом Lagurus lagurus. Специальные исследования показали, что пеструшки плейстоценовых популяций морфологически отличаются от современных.
Серые лемминги трибы Lemmini представлены единичными находками по всей толще отложений от уровня 10 горизонта 22.2 до горизонта 11.1. При этом только в отложениях слоя 20 количество останков лемминга достигает 3,2%. В остальных случаях оно не превышает 1 %.
Узкочерепная полевка Stenocranius gregalis - один из массовых видов плейстоценовых отложений. В каждом из слоев количество Mj узкочерепной полевки превышает количество Mj любого другого вида полевок в 2 - 3 раза. В среднем количество ее останков колеблется от 7 до 10%: в отложениях горизонтов 13.4 и 13.1 оно превышает 14%, а в горизонте 12.2 -15%. Относительная численность узкочерепной полевки возрастает вверх по разрезу до горизонта 12.2, а затем начинает снижаться. В целом похожую тенденцию отражают сборы 1994 г., хотя полного совпадения отдельных пиков по материалам разных квадратов не наблюдается.
Полевка-экономка Microtus oeconomus отмечена по всей толще плейстоценовых осадков. Однако ее численность редко достигает 3%. Только в отложениях горизонта 14.3 количество этой полевки возрастает до 5,9%. Правда в этом горизонте обнаружено небольшое количество костного материала и данный показатель может иметь случайный характер.
Северосибирская полевка Microtus hyperboreus -редкий представитель тафоценоза. Останки этой полевки встречаются спорадически, главным образом в средней и верхней частях разреза.
Темная полевка Microtus agrestis - редко встречается в отложениях плейстоценовой толщи. Отдельные зубы отмечены в слое 19, а также в горизонтах 14.2,14.1,13.4,12.1 и 11.4. Темная полевка - эволюционно молодой вид. Его выделение из общего ствола серых полевок Microtini произошло в первой половине среднего неоплейстоцена. Современные морфологические черты этот вид приобрел во второй половине среднего неоплейстоцена.
Обыкновенная полевка Microtus arvalis тоже редка в плейстоценовых отложениях пещеры. Ее малочисленные останки зафиксированы от слоя 21 до горизонта 11.2.
Водяная полевка Arvicola sp. - редкий вид плейстоценовых осадков. Однако ее останки встречаются почти по всему разрезу: от уровня 5 горизонта 22.1 до горизонта 9.3. Водяная полевка представлена эволю-ционно продвинутой формой, близкой современной Arvicola terrestris.
Алтайский цокор Myospalax myospalax является одним из самых характерных элементов тафоценоза плейстоценовых отложений. Его останки встречаются практически на всех горизонтах разреза. Их относительная численность колеблется от 5 до 7%, а в слое 20 достигает 13%. Останки цокора являются важным палео-климатическим показателем. Они свидетельствуют об относительной стабильности природной среды на протяжении всего периода накопления плейстоценовых осадков. Примечательно, что в двух колонках 1994 и 1995 гг. фиксируется одинаковое падение численности цокора в осадках горизонта 22.3, а также в отложениях слоя 17 и нижней части слоя 14.
Пищуха Ochotona sp. представляет немногочисленный, но постоянно встречающийся компонент тафоценоза. Относительная численность пищухи, как правило, не достигает 3%. Однако на уровне 3 горизонта 22.1 она превышает 4,8%, а в слое 20 достигает 5,2%. В отложениях пещеры присутствует два вида пищухи: алтайская Ochotona alpina и степная О. pusilla. Первый вид и сегодня обитает в бассейне Ануя. Горный Алтай является одним из районов его возникновения и формирования в процессе эволюционного становления. Степная пищуха - типичный обитатель равнинных степных ландшафтов Евразии. Его проникновение в бассейн Ануя могло произойти только в специфических природных условиях, т.е. при увеличении доли степных сообществ. Останки двух видов пищух подчеркивают своеобразие и пестроту плейстоценовых ландшафтов в окрестностях пещеры.
Заяц Lepus sp. редко встречается в костном материале мелкой фауны. Однако разрозненные зубы этого животного отмечены в плейстоценовой толще от уровня 8 горизонта 22.2 до горизонта 9.3. При этом их относительное количество не превышает 1-1,5%. Судя по размеру фрагментов и целых зубов, основная их часть принадлежит зайцу-толаю.
Мелкие куньи рода ласки Mustela sp. в небольшом количестве присутствуют почти по всему разрезу. В связи с фрагментарностью и малым количеством определимого материала вопрос о механизме их попадания в пещерные отложения пока остается открытым.
Птицы Aves. Кости птиц и скорлупа птичьих яиц часто встречаются в отложениях пещеры. В среднем их относительная численность составляет 5 - 7%, возрастая до 19,7% в слое 20. В толще слоя 22 число останков птиц резко снижается до 1,5- 2%. Возможно, это связано с большей залесенностью и закрытостью биотопов в эпоху накопления слоя 22. Подобная тенденция отмечена и в сборах 1994 г., хотя там она выражена менее отчетливо.
Рептилии Reptilia. Редко встречаются в плейстоценовых отложениях пещеры. Они представлены разрозненными фрагментами челюстей ящериц преимущественно в средней и верхней частях разреза.
Амфибии Amphibia. Кости конечностей лягушек спорадично встречаются по всему разрезу. Наибольшее количество останков амфибий отмечено в стратиграфическом интервале горизонтов 19.3 - 13.2.
Рыбы Pisces. Один из наиболее многочисленных компонентов тафоценоза пещеры. В среднем останки рыб составляют 10 - 12%. В отложениях горизонтов 21.1 и 9.3 их количество достигает более 28%. В пределах слоя 22 относительная численность костных остатков рыб снижается до 3 - 5%.
Динамика численности отдельных таксонов по сборам 1995 г. позволяет проследить общие закономерности изменения сообщества мелких млекопитающих за время накопления толщи плейстоценовых осадков Денисовой пещеры. В целом облик микротериофауны определяют типичные обитатели сухих и высокогорных степей — Stenocranius gregalis и Alticola strelzovi. Третьим важным компонентом является степная пеструшка Lagurus lagurus. От нижнего уровня слоя 22 до кровли плейстоценовой толщи встречаются останки характерных представителей лесных биотопов - полевок рода Clethrionomys. Практически на всех уровнях разреза отмечены останки Asioscalops altaica и Myospalax myospalax. Оба вида - специализированные землерои, которые не могут существовать при глубоком промерзании почвы. К видам, встречающимся в небольших количествах, но по всему разрезу, относятся Cricetulus barabensis и Microtus oeconomus. Первый из них обитатель степей, второй - пойменных биотопов.
Высокая численность в составе фауны степных и нивальных видов предполагает существование значительных участков открытых ландшафтов в период накопления плейстоценовой толщи. Вместе с тем присутствие останков Clethrionomys, единичные находки Sciurus, Pteromys volans, Eutamias sibiricus свидетельствуют о постоянном наличии лесных массивов.
Обитатели леса наиболее многочисленны в слое 22, где доля Clethrionomys составляет на разных уровнях от 5 до 11% общего состава. Удельный вес Alticola колеблется от 3,8 до 15%. Достаточно заметным элементом сообщества является Stenocranius - 3 - 5%. Интересно отметить обилие в этом слое костных остатков летучих мышей. На отдельных уровнях останки Chiroptera составляют более 60%. В слоях 20 - 9 их показатель не превышает 3 - 6%, что, очевидно, свидетельствует о возрастании фактора беспокойства со стороны человека. В целом накопление слоя 22, видимо, происходило в период широкого развития лесной растительности при умеренных климатических режимах.
В отложениях слоя 21 численность Clethrionomys существенно не меняется, удельный вес Alticola увеличивается до 16,4%, а доля Stenocranius составляет 4,3%. С этого уровня в ископаемой фауне начинают постоянно встречаться костные остатки Lagurus lagurus.
Своеобразные изменения в составе мелких млекопитающих отмечены в слое 20. Здесь резко сокращается численность Clethrionomys, снижается до минимума количество Alticola и Lagurus, а доля Stenocranius gregalis возрастает до 7,2%. Для этого слоя характерны наиболее высокие значения Myospalax myospalax (13%) и Asioscalops altaica (5,2%). Видимо, в период накопления слоя 20 происходило одновременное сокращение лесных массивов и участков горно-степных петрофильных ассоциаций. В это время наибольшего развития достигли лугово-степные ассоциации и ли-ственнично-березовые леса. Это была эпоха лугового и лесного разнотравья с минимальным участием злаков. Климат отличался теплыми зимами и дождливым летним периодом.
Выше по разрезу численность основных групп мелких млекопитающих восстанавливается в прежних соотношениях и сохраняет относительную стабильность до кровли слоя 14. В отложениях слоев 19-14 доля Clethrionomys варьирует в пределах 5 - 7%, Alticola -9 - 20%, Stenocranius - 5 - 10%, Lagurus - 3 - 5%.
В отложениях горизонта 13.2 отмечено резкое падение численности Clethrionomys и Alticola, количество Stenocranius gregalis снижается до 3,6%, исчезает Asioscalops, Lagurus и Microtus oeconomus. Одновременно увеличивается численность Lemmini. Вероятно, формирование этого горизонта связано с эпохой господства горно-степных петрофильных и нивальных злаково-осоковых ассоциаций в условиях относительно холодного и сухого климата.
Общий облик тафоценоза восстанавливается на уровне горизонта 13.1. Здесь численность Clethrionomys достигает 6,5%, Alticola - 13,3%, Stenocranius gregalis - 14,4%, появляются Asioscalops и Microlus оесопотш. Однако такая стабилизация природной обстановки была кратковременной.
В интервале слоев 12-9 наблюдается заметная перестройка в составе плейстоценовой биоты. На фоне относительно высокого удельного веса Alticola (12-17%) и Stenocranius (8 - 15%) удельный вес Clethrionomys снижается до 1 - 2%, a Lagurus - резко возрастает, достигая своего максимума (17,8%) в отложениях горизонта 9.1. Эти данные свидетельствуют о значительных изменениях ландшафтной обстановки. Очевидно, накопление осадков этой части разреза проходило в период сокращения участков леса и увеличения площади открытых биотопов, занятых в основном горно-степными петрофильными ассоциациями. Особенно ярко эта тенденция выражена в отложениях слоя 9.
Диаграммы состава мелких позвоночных имеют незавершенный характер. Нарастание остепнения, ухудшение климатической обстановки не достигают здесь своего максимума. Исходный материал не демонстрирует перехода от плейстоценовой биоты к го-лоценовой. Эти данные указывают на отсутствие в толще пещерных отложений осадков финального этапа плейстоцена.
Все мелкие млекопитающие, представленные в та-фоценозе пещеры, по своей морфологии мало отличаются от аналогичных современных таксонов. Вместе с тем данное сообщество по структуре тафоценоза, количественному соотношению входящих в его состав видов, напротив, принципиально отличается от современных сообществ мелких млекопитающих бассейна Ануя.
5.6. Орнитофауна палеолита Северо-Западного Алтая
Костные остатки птиц, по определению А.В. Пантелеева [2002], принадлежат 66 таксонам: Anas querquedula, Anas clypeata, Aythya fuligula, Anatidae indet., Aquila sp., Buteo lagopus, Aegypius monachus, Circus cyaneus, Circus sp., Accipitridae indet., Cerchneis tinnunculus, Hypotriorchis subbuteo, Tetraogallus altaicus, Perdix perdix/dauuricae, Coturnix coturnix, Lagopus mutus, Lagopus lagopus, Lagopus sp., Tetrao urogallus, Lyrurus tetrix, Galliformes indet., Porzana porzana, Tringa glareola, Charadriiformes indet., Athene noctua, Asio flammeus, Strigidae indet., Apus apus, Dendrocopos minor, Picoides tridactylus, Melanocorypha yeltoniensis, Calandrella rufescens, Eremophila alpestris, Alaudidae indet., Hirundo rustica, Cercopis daurica, Anthus trivialis, Anthus hodgsoni, Anthus cf. gustavi, Cinclus cinclus, Tarsiger cyanurus, Phoenicurus ochruros.
Oenanthe oenanthe, Turdus ruficollis, Turdus sp., Acrocephalus sp., Sylviidae indet., Emberiza da, Emberiza hortulana, Emberiza aureola, Plectrophenax nivulis, Leucosticte arctoa, Uragus sibiricus, Carpodacus erythrinus, Pinicola enucleator, Pyrrhula pyrrhula, Fringillidae indet., Montifringilla nivalis, Sturnus vulgaris, Pica pica, Pyrrhocorax pyrrhocorax, Corvus monedula/dauuricus, Corvus cornix/corone, Corvidae indet., Passeriformes indet., Aves indet.
В целом плейстоценовая орнитофауна имеет аль-пийско-степной облик (табл. 4): преобладают останки высокогорных видов, среди которых наиболее обильны Leucosticte arctoa (24,0%), Lagopus lagopus (23,1%), Plectrophenax nivalis (5,7%), а также Eremophila alpestris (5,5%), обитающий в степях от низин до высокогорий. Лесных птиц мало как по числу видов, так и по количеству останков.
В отложениях слоя 22 доминируют (77,6%) останки птиц альпийской и гольцовой зон, доля степных форм составляет 8,4%, на таежные виды приходится 5,6%, птицам скал и опушки леса принадлежит соответственно 3,5 и 4,9% костей.
В средней части плейстоценовой толщи наиболее обеспечены орнитофауной и сопоставимы между собой материалы слоев 20,19,12 и 11. Останки птиц различных горных биотопов представлены в следующем соотношении: 1) высокогорные виды - слой 20 - 44,4%, слой 19 - 55,6%, слой 12 - 56,8%, слой 11 - 57,8%; 2) птицы скал и открытых крутых склонов - слой 20 -2,8%, слой 19 - 5,5%, слой 12 - 6,8%, слой 11 - 3,9%; 3) степные виды - слой 20 -16,7%, слой 19-13,9%, слой 12 - 9,1 %, слой 11 - 8,8%; 4) таежные виды - слой 20 -33,3%, слой 19-11,1%, слой 12-9,1%, слой 11 -9,8%; 5) птицы опушки леса и горных лугов - слой 20 - 2,8%, слой 19 - 13,9%, слой 12 - 11,4%, слой 11 - 9,8%; 6) птицы лиственного леса - слои 20, 19 - 0%, слой 12 -4,5%, слой 11 - 4,9%; 7) околоводные виды - слои 20, 19 - 0%, слой 12 - 2,3%, слой 11 - 4,9%.
В верхней части разреза (слой 9) значение альпийской группы практически не изменилось - 58,3%. Вместе с тем заметно возросла доля птиц - обитателей скал - 25,0%. Показатель таежных видов сократился до 4,2%, хотя костные остатки птиц опушки леса составили 12,5%. Птицы лиственных лесов, степей и околоводных биотопов в этом слое не отмечены.
 Табл.
61. Хроностратиграфия палеолита
долины Ануя (в
книге эта важнейшая таблица, без которой мало что можно тут понять, помещена
почему-то в самом конце 10 главы, где ее довольно трудно найти. Я переместил ее
сюда для удобства читателей - А.М.).
Табл.
61. Хроностратиграфия палеолита
долины Ануя (в
книге эта важнейшая таблица, без которой мало что можно тут понять, помещена
почему-то в самом конце 10 главы, где ее довольно трудно найти. Я переместил ее
сюда для удобства читателей - А.М.).
Первоначальными данными для хронологического членения разреза послужили даты, полученные радиоуглеродным методом по гуминовым кислотам и кости: слой 11 - более 37 235 лет (СОАН-2504); слой 21 -39 360 ± 1 310 лет (СОАН-2499) и более 34 700 лет (СОАН-2488). В то же время другие данные хроно-стратиграфии (археологические, литолого-стратигра-фические и биостратиграфические) свидетельствовали о более широких возрастных пределах для плейстоценовых отложений пещеры.
На следующем этапе исследований была проведена оценка возраста плейстоценовой толщи физическими методами датирования. Для отложений средней и нижней частей разреза О.А. Куликов получил серию РТЛ-дат: горизонт 14.1 - 69 ± 17 тыс. л.н. (РТЛ-611); слой 21 - 155 ± 31 тыс. л.н. (РТЛ-546); горизонт 22.1 -171 ± 43 тыс. л.н. (РТЛ-737) и 182 ± 45 тыс. л.н. (РТЛ-738), верхняя часть горизонта 22.2 - 223 ±55 тыс. л.н. (РТЛ-739) и 224 ± 45 тыс. л.н. (РТЛ-547), нижняя часть горизонта 22.2 - 282 ± 56 тыс. л.н. (РТЛ-548).
Результаты РТЛ-датирования нижней части разреза хорошо согласуются с их палеомагнитной характеристикой. По данным З.Н. Гнибиденко, на фоне прямой полярности плейстоценовых отложений, свойственной эпохе Брюнес, в осадках слоя 22 установлены два экскурса геомагнитного поля: Бива I (176 - 220 тыс. лет) для горизонта 22.1 и Бива II (266 -300 тыс. лет) для горизонта 22.2 [Деревянко, Гнибиденко, Шуньков, 1998].
Сначала средненеоплейстоценовый возраст базаль-ных отложений Денисовой пещеры был воспринят с большой долей осторожности [Археология..., 1998; Шуньков, Агаджанян, 2000; Paleolithic..., 2001]. Вместе с тем развитие ранней стадии среднего палеолита в хронологических рамках среднего неоплейстоцена было установлено в других классических районах изучения палеолита Евразии. Так, на территории Западной и Центральной Европы в начале рисского времени наряду с типичными комплексами ашеля отмечено развитие пре- и раннемустьерских индустрии без ашельских бифасов, с развитыми орудийными формами на отщепах [Bosinski, 1982; Roe, 1982; Tuffreau, 1982]. На Ближнем Востоке по данным ТЛ-датирования обожженных кремней из культурно-стратиграфической колонки пещеры Табун хронологические рамки мугаранской традиции (слои Ed- Еа), включавшей ябрудьен, установлены в пределах 270 - 350 тыс. лет, а ранний этап развития леваллуа-мустьерских индустрии (слой D) относится к интервалу 244 - 263 тыс. лет [Jelinek, 1992; Bar-Yosef, 1995; Mercier, Valladas and Valladas, 1995; Mercier etal., 1995]. На этом фоне хронометрические показатели нижних среднепалеолитических слоев Денисовой пещеры выглядят вполне достоверно.
В целом РТЛ-даты пещерных отложений не противоречат общему стратиграфическому контексту плейстоценовой толщи и могут рассматриваться в качестве хронологических отметок для определения геологического возраста выделенных здесь климатостра-тиграфических подразделений. При этом следует подчеркнуть, что литологические и биостратиграфические характеристики пещерного осадка указывают на весьма значительную суммарную продолжительность седиментационных перерывов, т.е. на относительную неполноту летописи плейстоценовых отложений пещеры.
Согласно результатам комплекса методов абсолютной и относительной геохронологии, в качестве рабочей схемы принята следующая хронологическая последовательность для отложений Денисовой пещеры.
Палеогеографические данные условий формирования наиболее древних отложений (горизонт 22.3 и нижняя часть горизонта 22.2) отражают благоприятную климатическую обстановку с достаточно теплым и умеренно влажным климатом. Согласно РТЛ-дате 282 ± 56 тыс. л.н., возраст этих отложений находится у верхней хронологической границы тобольского (соуськанихинского, М - R) межгляциала среднего неоплейстоцена, что, возможно, соответствует возрасту стадии 9 морской изотопной стратиграфии [Карабанов и др.,2001].
Для эпохи относительного похолодания и общего ухудшения природной обстановки, зафиксированной в осадках верхней части горизонта 22.2, получено два хронологических измерения - 223 ± 55 тыс. лет и 224 ± 45 тыс. лет. Возрастные оценки этих отложений соответствуют возрасту самаровского (ештыккольского, r1) ледникового горизонта, который, возможно, коррелируется с изотопной стадией 8.
Период накопления горизонта 22.1 отличался стабильной природной обстановкой с небольшими колебаниями влажности и температуры, которые не выходили за рамки параметров климата межледниковья.
Показатели возраста этих отложений -171 ± 43 тыс. лет и 182 ± 45 тыс. лет - находятся в пределах ширтинского (чаганузунского, R,,) межгляциала, который, возможно, соответствует изотопной стадии 7.
Слой 21 сформировался в условиях относительно холодного и умеренно влажного климата на заключительной стадии среднего неоплейстоцена. Судя по РТЛ-дате 155 ± 31 тыс. л.н., эта климатическая фаза относится к тазовскому (чуйскому, Rm) ледниковому периоду (изотопная стадия 6).
Осадки слоя 20 и нижней части слоя 19 соответствуют этапу теплого и относительно сухого климата, для которого характерно широкое развитие сосново-березовых лесов с участием широколиственных видов. Согласно стратиграфической позиции в толще пещерных отложений, накопление этих осадков происходило, скорее всего, в начале верхнего неоплейстоцена -в казанцевский (куэхтанарский, R-W) климатический оптимум (изотопная под стадия 5е).
Отложения слоев 19 (верхняя часть), 17 и 14 формировались после седиментационного перерыва в условиях неустойчивой климатической обстановки, когда в составе лесных массивов заметно увеличилась доля темнохвоиных пород. РТЛ-возраст кровли слоя 14 -69 ± 17 тыс. л.н. позволяет сопоставить этот период в целом с первой половиной ермаковского (чибитского, W,) времени (изотопные подстадии 5а- 5б).
На заключительном этапе ермаковского времени (изотопная стадия 4), в эпоху накопления слоев 13 и 12 отмечено общее ухудшение климатической обстановки. Биостратиграфические показатели этого интервала разреза фиксируют неоднократную перестройку растительных ассоциаций в условиях чередования климата холодного сухого (резкое сокращение лесных биотопов), холодного влажного (активизация темнохвоиных лесов) и более сухого переходного (возобновление березовых лесов с участием ольхи и лещины).
Эпоха образования слоя 11, судя по заметному изменению ландшафтно-климатической обстановки, относится к следующей палеогеографической фазе верхнего неоплейстоцена. Для нее характерно широкое распространение темнохвоиных лесов и луговых сообществ в условиях относительно прохладного и влажного климата. Радиоуглеродный возраст этого слоя составляет более 37 235 лет, что вполне соответствует первой половине каргинского (бельтирского, WII) времени (изотопная стадия 3).
Формирование верхней части плейстоценовой толщи происходило после длительного седиментационного перерыва, отмеченного в структуре осадка слоя 10. Экологический облик растительности и животного мира в эпоху накопления слоя 9 свидетельствует о максимальном развитии нивальных и степных биоценозов в холодной и сухой климатической обстановке сартанского (аккемского, WIII) времени (изотопная стадия 2).
5.10. Антропологические материалы палеолита Северо-Западного Алтая
Антропологические материалы представлены двумя одонтологическими образцами из коллекции 1984 г. По определению Е.Г. Шлаковой [2001], в отложениях горизонта 22.1 обнаружен левый нижний второй молочный моляр т2 ребенка 7-8 лет, а в отложениях слоя 12 - левый верхний медиальный резец I1 взрослого субъекта. Этот материал имеет чрезвычайно важное значение в изучении вопросов последовательности заселения территории Горного Алтая представителями рода Homo. Поэтому образцы зубов из Денисовой пещеры исследовались несколькими специалистами, в результате чего были получены три различные интерпретации таксономической принадлежности одонтологического материала.
Первоначально, по идентификации К. Тернера, предполагалось, что зубы ископаемых гоминидов из Денисовой пещеры обладают рядом хорошо выраженных черт неандертальского одонтологического комплекса, тяготеющего к представителям европейских, а не азиатских групп Homo sapiens neandertalensis [Turner, 1988,1990а, б].
В дальнейшем исследование зубов провел В.П. Алексеев. Единичность образцов и состояние, соответствующее возрасту субъектов, к которым они относились, не позволили ему оценить перспективность находок в плане их таксономической принадлежности. По мнению В.П. Алексеева, морфологические особенности этих зубов не могут однозначно быть сопоставлены с комплексом неандертальской группы гоминидов и допускают их связь с ископаемым человеком современного физического типа [Alekseev, 1998].
Наличие двух альтернативных интерпретаций послужило поводом для проведения Е.Г. Шлаковой детального анализа сохранившихся на зубах морфологических особенностей. По совокупности метрических показателей и описательных признаков она установила, что, несмотря на отдельные архаичные черты, одонтологический материал Денисовой пещеры принадлежит, скорее всего, представителям ископаемого человека современного физического типа - ранним Homo sapiens sapiens [Шпакова, 2001].
Мнение К. Тернера о таксономической принадлежности одонтологического материала из пещер Горного Алтая (в совокупности алтайские материалы включают образцы зубов из пещер Денисова и Окладникова) с момента его опубликования широко цитируется в российских и зарубежных исследованиях по антропогенезу, например, в таком авторитетном издании, как "Энциклопедия эволюции человека и предыстории" [Encyclopedia.. .,2000]. Однако в настоящее время более аргументированным выглядит заключение Е.Г. Шлаковой. Прежде всего она уточнила позицию расположения на челюсти молочного моляра и привела бесспорные доказательства, что этот зуб является вторым моляром левой стороны нижней челюсти. К. Тернер определил его как первый правый моляр. Размеры второго молочного моляра у гоминидов, как правило, превышают размеры первого и выбиваются из вариационного ряда метрических характеристик первых моляров, сближаясь с зубами неандертальских форм. При помещении этих размеров в вариационный ряд вторых моляров они абсолютно сопоставимы с параметрами зубов ископаемых людей современного физического типа. Кроме того, соотношение горизонтальных диаметров коронки исследованного зуба отражает редукцию вестибулолингвального диаметра, сопоставимую с этой тенденцией у представителей ископаемых форм Homo sapiens sapiens.
Форма лингвальной поверхности медиального резца взрослого субъекта также интерпретируется Е.Г. Шпаковой как нехарактерная для пренеандертальских и неандертальских групп, в которых не наблюдается сочетания мощного мезиального краевого гребня с отсутствием дистального, как это имеет место на резце из Денисовой пещеры.
Хорошая сохранность эмали резца и отсутствие на ней следов гипоплазии свидетельствуют, по мнению Е.Г. Шпаковой, о достаточно стабильном биологическом развитии субъекта, которому он принадлежал. Согласно этим признакам, можно предположить, что палеолитические обитатели Денисовой пещеры существовали в относительно комфортных условиях окружающей среды, без сильных физиологических стрессов, которые могли быть связаны с длительным голоданием или хроническими болезнями [Шпакова, Деревянко, 2000].
Перечисленные аргументы ставят под сомнение первоначальное определение К. Тернера и позволяют считать более обоснованным заключение Е.Г. Шпаковой о принадлежности одонтологических материалов из Денисовой пещеры представителям ископаемого человека современного физического типа - ранним Homo sapiens sapiens. Окончательное решение этого вопроса должно опираться на данные палеогенетического анализа.